
| What We Study |

|
The goal of the PHACE experiment is to understand how elevated CO2 and warming influence plants and soils. Specifically, we hope to learn how these changes influence the following key components of mixed-grass prairie:
|
Water controls most processes in the semiarid mixed-grass prairie. Elevated CO2 can decrease transpiration and increase water use efficiency and water availability. Warming often increases both evaporation and transpiration.
Nitrogen often limits plant productivity and quality, and can become increasingly limiting with elevated CO2.
Plant productivity and quality determine the ability of mixed-grass prairie to support grazing by livestock and wildlife, which are in turn key to the sustainability of agriculture and biological diversity.
Carbon cycling determines whether mixed-grass prairie sequesters or releases CO2, and therefore whether it contributes to or mitigates climate change. As the substrate for heterotrophic microbes, carbon also affects mineralization and immobilization of other nutrients.
Plant communities both respond to and influence biogeochemical cycling. In mixed-grass prairie, warm-season and cool-season grasses comprise the majority of plant biomass, while forbs comprise most of plant diversity. Legumes and sub-shrubs are less common, but can respond strongly to elevated CO2.
Invasive plants reduce forage production and biological diversity, and can thrive following increases in resource availability, including CO2 and water. PHACE includes several sub-experiments designed to test relative responses of native and invasive plants.
Microbes mediate biogeochemical cycling, and the structure of microbial communities can control both carbon cycling and nutrient availability to plants. |
 |

WHAT WE HAVE LEARNED
1. In this semi-arid grassland, elevated CO2 effects on soil water content counteract the desiccating effect of warmer temperatures. As anticipated, warming induced soil drying, but elevated CO2 increased soil water content more than expected, by increasing plant water use efficiency. Surprisingly, the combination of elevated CO2 and elevated temperature resulted in no difference in soil water content between these "future" plots and control plots under present-day CO2 and temperature levels (Figure 1).

Figure 1. Responses of soil water content (SWC) to CO2 & warming.Average and s.e.m. (error bars) of volumetric SWC (5-25 cm depth) for plots exposed to present-day ambient CO2 and temperature (ct), 1.5/3o C day/night warming (cT),600 p.p.m.v. CO2 (Ct), and 600 p.p.m.v. CO2 and 1.5/3o C day/night warming (CT) (five replications per treatment). Data are averaged over days of year (DOY) 100-200, the early- to mid-growing season when soil water most limits productivity. Precipitation amounts for this same period are also presented. Significance (P ≤ 0.05) for main effects and year are given in the figure (Figure 1 from Morgan et al. 2011).
These soil water effects influenced plant biomass responses. Higher CO2 increased plant biomass, especially in dry years when water savings were most important to growth (2006-2008). Warming alone did not significantly affect total aboveground biomass, likely because potential increases in plant growth were limited by lower soil water. There were no net decreases in plant growth between present-day and future conditions (elevated CO2 and temperature).
The effect of higher CO2 on soil water appears to be the dominant driver of plant biomass responses in semi-arid grasslands. Relative to plots at ambient CO2, the increase in aboveground biomass at elevated CO2 is stronger at a more negative early-season soil matric potential (i.e., drier soil), both at PHACE and an earlier CO2 enrichment experiment (Morgan et al. 2007). This relationship from two different ecosystems suggests that CO2 will increase plant productivity when plants are actively growing but water-limited (Figure 2).

Figure 2. Response of biomass enhancement ratio to soil matric potential.Effects of early-season (DOY 100-200) rooting zone soil matric potential (ψm) on biomass enhancement ratio, the ratio of mid-July harvested above-ground plant biomass in CO2-enriched plots divided by plant biomass from ambient CO2 plots. Data are from the PHACE experiment (4 years' data, n = 4), and from a previous open top chamber CO2 enrichment experiment (5 years' data, n = 5) conducted on Colorado shortgrass steppe. For further details, see Supplementary Appendix I, soil water conversions (Figure 3 from Morgan et al. 2011).
For more results, see: Morgan, J.A. et al. 2011. C4 grasses prosper as carbon dioxide eliminates dessication in warmed semi-arid grassland. Nature 476: 202-206. and Morgan, J. A., et al. 2007. Carbon dioxide enrichment alters plant community structure and accelerates shrub growth in the shortgrass steppe. Proc. Natl Acad. Sci. USA 104, 14724-14729.
2. Elevated CO2 and warming have contrasting effects on N availability as well as moisture and productivity (Figure 3). Under elevated CO2, soil inorganic N decreased, likely due to increased microbial N immobilization. The CO2-induced increase in soil moisture facilitated higher N uptake by microbes, but did not affect plant N pool sizes. In contrast, warming increased soil inorganic N and plant N. Direct effects of warming on net N mineralization appeared to be more important than a warming-induced decrease in soil moisture.

Figure 3. Soil inorganic nitrogen (N) availability (NH4+ + NO3-) at 2-7.6 cm soil depth in 2006 averaged by CO2, and in 2007 and 2008 averaged by CO2, warming, and irrigation treatments.Treatments: ct, ambient CO2 and ambient temperature; cT, ambient CO2 and elevated temperature; Ct, elevated CO2 and ambient temperature; CT, elevated CO2 and elevated temperature; ct-i, ambient CO2 and ambient temperature, but irrigated. Soil inorganic N availability is expressed in μg N 10 cm-2 resin membrane area of the plant root simulator (PRS) probes per month incubation time. Error bars indicate ± 1 SE. ANOVA P-values are reported when P < 0.05 (in bold) or P < 0.1 (in italics) (Figure 2 from Dijkstra et al. 2010).
For more results, see: Dijkstra, F.A. et al. 2010. Contrasting effects of elevated CO2 and warming on nitrogen cycling in a semiarid grassland. New Phytologist 187: 426-437. and Carillo et al. 2012. Controls over soil nitrogen pools in a semiarid grassland under elevated CO2 and warming. Ecosystems 15: 761-774.
3. Wetter soil conditions under elevated CO2 increased P availability to plants and microbes relative to that of N, while drier conditions with warming reduced P availability relative to N (Figure 4). Soil moisture exerts an important control on inorganic P supply from desorption and dissolution reactions. This means that despite the fact that warming may alleviate N limitation under elevated CO2, warming and drought can exacerbate P limitation on growth and microbial activity.

Figure 4. N:P ratios in green (a, e) and senesced (b) plant tissue, microbes (c) and on Plant Root Simulator resin probes (PRS probes) (d) in response to elevated CO2, warming and irrigation (ct, ambient CO2 and temperature; cT, ambient CO2 and 1.5/3o C day/night warming; Ct, 600 ppmv CO2 and ambient temperature; CT, 600 ppmv CO2 and 1.5/3o C day/night warming; ct-i, ambient CO2 and temperature with 60 mm yr-1 irrigation). Green plant N:P ratios are species-weighted averages. Error bars represent 1SE. ANOVA P values are reported when P < 0.05 (in bold) or P < 0.1 (in italics) (Figure 1 from Dijkstra et al. 2012).
For more results, see: Dijkstra, F.A. et al. 2012. Climate change alters stoichiometry of phosphorus and nitrogen in a semiarid grassland. New Phytologist 196:807-815.
4. The PHACE plots were warmed by a free-air, feedback controlled infra-red heater system with target warming of 1.5/3.0 C day/night. We assessed how well the system maintained these target temperatures. Thermocouple temperature sensors were placed on leaves, in the soil, and above and within the plant canopy to monitor the actual warming at ecologically important zones.

Figure 5. The infra-red heater warming system worked very well. The leaf and soil temperatures were warmed very close to the day/night target warming of 1.5/3.0o C (note the green differential temperature bars). The warming system also performed well under very windy Wyoming conditions, and during both summer and winter.
For more results on the performance of the infra-red warming system see LeCain et al. 2015. Microclimate Performance of a Free-Air Warming and CO2 Enrichment Experiment in Windy Wyoming, USA. PLoS ONE. 10(2): e0116834. doi:10.1371/journal.pone.0116834
The data from the manuscript are available here: TFACE Solstice Power Use,Temperature Data, Solstice temperatures, Wind speed vs. Temp differences.
5. After seven years of the PHACE experiment (2007-2013), elevated CO2 and warming together increasingly favored cool-season (C3) graminoids, reversing an initial shift toward warm-season (C4) grasses (primarily, blue grama, Bouteloua gracilis) (panel b, outlined below in Figure 6). Forbs declined in response to warming alone. Plant community composition changed progressively from the beginning to the end of the study, while plant biomass was regulated by year to year differences in precipitation.
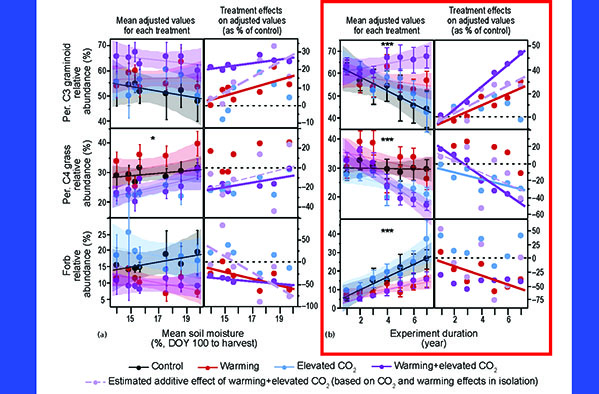
Figure 6. Influence of (a) soil moisture and (b) experiment duration on plant community composition and treatment effects on plant community composition. For each treatment, solid lines show the linear trend between the temporal variable (soil moisture or experiment duration) and both: (1) adjusted values of and (2) proportional treatment effects on relative abundance for each plant functional group. Treatments shown: ambient CO2 and temperature (black); ambient CO2 and 1.5/3°C day/night warming (red); 600 ppmv CO2 and ambient temperature (blue); 600 ppmv CO2 and 1.5/3°C day/night warming (purple) (Figure 4 from Mueller et al. 2016).
Elevated CO2 and warming together consistently stimulated total plant biomass by about 25%, due in large part to increased root biomass (Figure 7). The combined effect of CO2 and warming on aboveground biomass was also positive, but less so in wet growing seasons than dry ones. Lower soil nitrogen availability in wet growing seasons may have limited the potential for aboveground biomass to increase in response to elevated CO2.
Figure 7. Treatment effects on aboveground, root, and total plant biomass across years, for ambient CO2 and temperature (black bars); ambient CO2 and 1.5/3°C day/night warming (red bars); 600 ppmv CO2 and ambient temperature (blue bars); 600 ppmv CO2 and 1.5/3°C day/night warming (purple bars) (taken from Figure 2, Mueller et al. 2016).
Soil water and nitrogen were sensitive to precipitation, CO2 and warming treatments, and changes in the plant community over time. Soil nitrate, in particular, became progressively more positive in the combined CO2-plus-warming treatment.
What does this mean for the future? Our results suggest that elevated CO2 and warming could increase below-ground carbon inputs in semiarid grasslands. Forage production may increase, although forage quality (as indicated here by nitrogen content) could decline. In addition, peak forage availability and quality may begin and end earlier in the growing season than at present, due to the shift from warm- to cool-season grasses, and the mixed-grass prairie may become less resilient to summer droughts and heat waves.
For more results, see: Mueller, K. E et al. 2016. Impacts of warming and elevated CO2 on a semi-arid grassland are non-additive, shift with precipitation, and reverse over time. Ecology Letters 19: 956–966.
6. Invasive plants are poised to take advantage of changes in the amount and timing of resources brought about by rising CO2 and warmer temperatures. The mechanisms behind these responses varied by species. Dalmatian toadflax (Linaria dalmatica) plants grew 13 times larger, produced 32 times as many seeds, and 7 times as many vegetative sprouts under elevated CO2. Toadflax benefitted from the direct effect of increased carbon and the indirect effect of increased soil water availability under higher CO2 (Figure 8).


Figure 8. Left: Dalmatian toadflax (Linaria dalmatica) plants tower above native grasses in a PHACE study plot under warming and elevated CO2. Right: Cumulative effects of elevated CO2 (c=ambient; C=600 ppmv) and warming (t=ambient; T=1.5/3°C day/night warming) on toadflax growth and reproduction per square meter (left panels), and per surviving plant in the second growing season, to exclude effects of first-year mortality (right panels) (Figure 1 from Blumenthal et al. 2013).
Warming more than tripled cheatgrass (Bromus tectorum) biomass and seed production by accelerating early spring growth. Warmer temperatures are likely to expand the phenological niche for cheatgrass, a winter annual weed, enhancing its ability to grow when native plants are inactive and competition for water and nutrients is least intense (Figure 9).

Figure 9. Cumulative effects of elevated CO2 (eCO2) and warming on cheatgrass (Bromus tectorum) aboveground biomass (solid bars) and seed mass (shaded bars). Note the ten-fold scale differences across both competition level and year. (Figure 1 from Blumenthal et al. 2016).
The increased risk of plant invasion highlights the need to develop adaptive management strategies, including targeted livestock grazing to suppress weeds and encourage competitive forage plants in mixed-grass prairie (Figure 10).

Figure 10. Warming and elevated CO2 may facilitate the spread of invasive plants such as Dalmatian toadflax (yellow flowering plant, left) and cheatgrass (red flowering grass, right) in mixed-grass prairie, displacing native plant communities.
For more results, see: Blumenthal, D. M. et al. 2013. Invasive forb benefits from water savings by native plants and carbon fertilization under elevated CO2 and warming. New Phytologist 200:1156-1165. and Blumenthal, D.M. et al. 2016. Cheatgrass is favored by warming but not CO2 enrichment in a semi-arid grassland. Global Change Biology 22: 3026-3038.